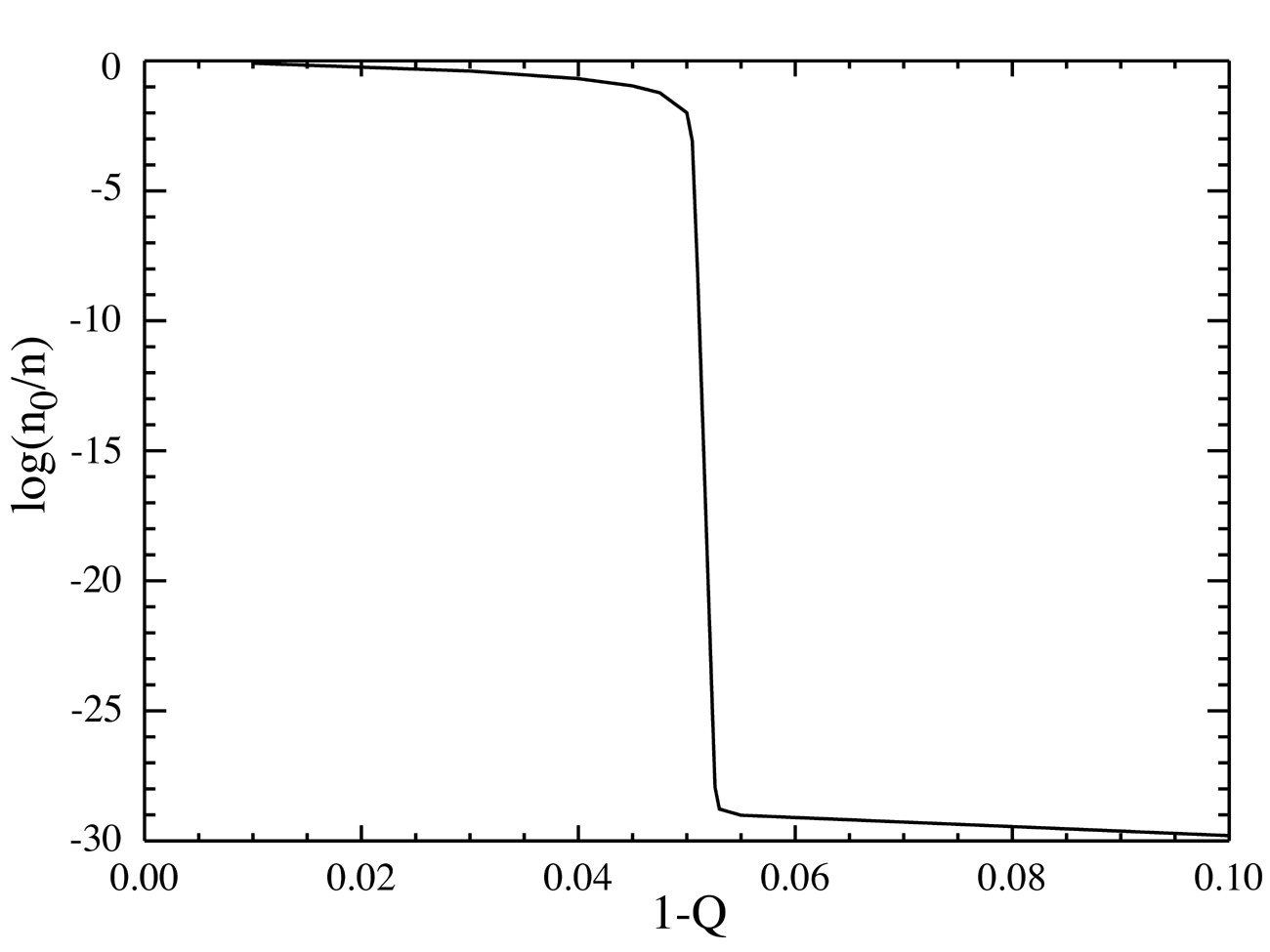
NOTE: the ordinate (Y-axis) is on a log scale, so the almost all-or none change in the concentration of the master is by a factor around 10^28 (ten to the twenty eight, a million times greater than Avogadro's number.
Here is part of the student's interpretation of my lecture on Eigen's model, and my response, in bold. More to follow....
"The Eigen model of molecular evolution allows us to make a connection between darwinian evolution and the origins of DNA/ RNA replication. Eigen's model works on the basis that in our universes early history, RNA were able to fold and function as catalysts for Polypeptide replication before DNA came into the picture. When DNA developed, the process changed slightly, including the creation of complimentary strands before the identical strand is produced. "
Not quite right. Eigen's model does not explicitly aim to model conditions on the early earth, nor to describe the special case of RNA replication. Indeed at the time the model was first developed (1970), the catalytic abiilities of RNA were not yet recognized. Eigen's model had a more general aim: to describe quantitatively a simplified and generalized model of polynucleotide (or indeed heteropolymer) replication. The first core feature is sequence copying - that the precise linear sequence of n monomers in a polymer made up of at least 2 different monomers (for convenience represented as 0 or 1) depends in a 1-to-1, 0 to 0 fashion on the precise linear sequence of another "template" sequence. All other specifics are left out. As I explained, we must consider the relative concentration of all possible 2^n possible sequences when each sequence is growing exponentially at different rates phi but also "killed" (eg by simple dilution) with equal probability (because every so often half the solution is thrown away, so as to keep the total number of molecules constant despite the ongoing replication.
So 2 key features of the model are sequence-specific replication and competition for resources (i.e. hi-energy nucleotides). The third crucial feature is the possibility of mistakes in copying individual monomers (eg bases) despite the high specificity of e.g Crick-Watson base pairing. This is a minimal model of Darwinian evolution at the molecular (not organismal) level. Eigen suspected, and was able to prove (both mathematically and experimentally - a nice combination) that if the error rate e exceeded a critical value approximately equal to 1/n, that Darwinian natural selection would stop. Or, applied to the Origin of Life problem, that since on the early earth replication (i.e. template-dependent copying) was probably rather inaccurate, Darwinian evolution could not start until the relevant catalyst (whatever it was) became accurate enough that e < 1/n. The whole point is that at least in the model there is a sharp dividing line (analogous to a phase transition) between purely chemical processes (eg low-accuracy copying) and Darwinian evolution (= Life) at a critical error rate ~ 1/n.
Note that while the model explicitly considers only sequences of fixed length n exactly the same outcome would be observed in a model with variable length n (for example, in the likely case that shorter sequences were more likely than longer ones). The model throws out all the interesting but basically irrelevant details to focus on the essence of the problem: point mutation.
No comments:
Post a Comment